O controle da respiração animal.
Santos, H.A.X.; Sousa, F.O.; Rezende, M.G.; Moraes, I.A. 2019.
Segundo Reece (2017) e Klein (2014) a respiração é o
processo pelo qual os animais obtêm e utilizam o oxigênio (O2) e
liberam o dióxido de Carbono (CO2). Juntamente com os rins, ela
desempenha um papel muito importante no equilíbrio acidobásico no organismo,
pois para que o pH do sangue seja mantido dentro da normalidade se faz
necessário o equilíbrio entre os gases da respiração (O2, CO2
e N2) e a adequada produção e eliminação de H+.
Klein (2014) informa que durante as atividades diárias, um
animal poder ter vários níveis de atividade e pode respirar ar de diferentes
composições e graus de pureza. Assim,
para permitir que o sistema respiratório responda a estes desafios, são
necessários mecanismos de controle que possam monitorar a composição química do
sangue, o esforço exercido pelos músculos respiratórios sobre os pulmões, e a
presença de matérias estranhos no trato respiratório. Segundo o autor, o controle
feito a fim de produzir um padrão respiratório que mantenha a troca gasosa
deverá estar integrado com outras atividades não respiratórias tais como a
termorregulação, vocalização, parto e a eructação.
Reece (2017) indica que a respiração dos animais deve ser
rigorosamente regulada para manter as concentrações de O2, CO2
e íons hidrogênio (H+) em níveis constantes, atendendo as
necessidades do corpo em diferentes condições. Se houver aumento na
concentração de H+ ou CO2, ou se a concentração de O2
diminuir, seus níveis serão normalizados por meio do aumento da ventilação. Em
contrapartida, se a concentração de H+ ou de CO2
diminuir, ou se houver aumento da concentração de O2, a ventilação pulmonar
irá diminuir.
É fácil notar que a respiração dos animais atende a um
padrão rítmico e automático com capacidade para ajustar às necessidades de
maior ou menor consumo de O2 ou eliminação de CO2. E isso
é possível pois existe um Centro Respiratório localizado no Sistema Nervosos Central
(SNC) que se encarrega de manter a respiração sob controle.
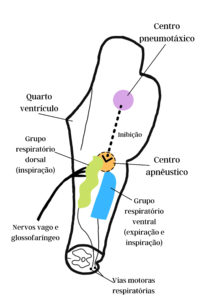
O
Centro respiratório.
Reece (2017) informa que o padrão rítmico da respiração e os ajustes que ocorrem são integrados nas partes do tronco encefálico conhecidas como centro respiratório. Segundo ele, diferentemente de muitos centros, o centro respiratório consiste em regiões no bulbo e na ponte associadas a funções especificas relacionadas com a respiração. Segundo o autor foram identificadas quatro regiões específicas: Grupo respiratório dorsal (GRD) – localizado na parte dorsal do bulbo; Grupo respiratório ventral (GRV) – localizado na parte ventral do bulbo; Centro pneumotáxico (CP) – localizado na porção rostral da ponte; Centro apnêustico (CA) – localizado na parte caudal da ponte.
Imagem 1 – Componentes do centro respiratório. Os centros apnêustico e pneumotáxico localizados na ponte e os grupos respiratórios ventral e dorsal localizados no bulbo. Adaptado de Dukes – Fisiologia dos animais domésticos. 13ª ed. Rio de Janeiro: Guanabara Koogan, 2016.
O grupo respiratório dorsal (GRD)
O GRD está principalmente associado à inspiração e gera o
ritmo básico da respiração. Ele recebe
neurônios aferentes dos pares dos nervos cranianos glossofaríngeo (IX) e vago (X)
e sua descarga (eferência) é transmitida pelo nervo frênico para o diafragma
proporcionando sua contração e a fase inspiratória do ciclo respiratório
(REECE, 2017). Os impulsos provenientes dos mecanorreceptores pulmonares
estimulados pela inflação dos pulmões chegam ao GRD por meio do nervo vago e
permitem a terminação da fase de inspiração, e os impulsos gerados nos quimiorreceptores
periféricos localizados por exemplo da pele, músculos e articulações chegam ao
GRD por meio do nervo vago e glossofaríngeo e estimulam a inspiração.
Reece (2017) afirma que GRD
dispõe de neurônios que atua tanto na inspiração quanto na expiração quando o
animal está em repouso e a expiração ocorre de modo passivo. No entanto durante
o exercício físico, quando a expiração se torna um processo ativo, os neurônios
expiratórios tornam-se ativos e garantem maior eficiência expiratória.
O grupo
respiratório ventral (GRV).
Segundo Reece (2017) o GRV
participa da fase de inspiração do ciclo respiratório. Esse grupo auxilia a
inspiração iniciada pelo GRD e ao mesmo tempo, inibe os neurônios expiratórios
do GRD durante a fase inspiratória do ciclo respiratório. É provável que os
neurônios inspiratórios do GRV também sejam mais ativos durante o exercício
físico
Centro pneumotáxico (CP)
O centro pneumotáxico (CP) age
inibindo a inspiração, e portanto, regula o volume inspiratório e a frequência
respiratória. A principal função do CP consiste em limitar a inspiração,
controlando, assim, a duração da fase de enchimento do ciclo respiratório
(REECE, 2017). O autor acrescenta que o sinal pneumotáxico, que desliga a
inspiração e encerra a fase de enchimento, pode ser forte ou fraco. O efeito de
um sinal forte consiste em aumentar a frequência respiratória, por meio da qual
tanto a inspiração quanto a expiração são encurtadas e estão acopladas a menor
volume corrente. O inverso é verdadeiro para um sinal fraco do CP.
Centro apnêustico (CA)
De todas as regiões do centro
respiratório, o centro apnêustico é a menos compreendida; em consequência, não
existe consenso sobre a função. Enquanto o CP está relacionado com o termino da
inspiração, acredita-se que o centro apnêustico esteja associado às inspirações
profundas (apneuse). Talvez as respirações complementares (suspiros) sejam
manifestações na atividade do centro apnêustico.
Controle integrado da respiração
Para que o controle da respiração seja efetivo é necessária a integração dos níveis central e periférico.
No nível central vamos
encontrar o centro respiratório na ponte e bulbo e córtex cerebral que permite
o controle voluntário da respiração por um determinado período até que as
células do centro respiratório escapem da inibição (Reece, 2017). No nível
central existem também quimiorreceptores que são áreas quimiossensíveis presentes
na superfície ventral do bulbo e extremamente sensíveis às mudanças na
concentração do íon hidrogênio que ocorrem pela difusão do CO2
pelo líquido cérebro-espinhal que está presente na região. O aumento de CO2
alveolar vai causar um aumento na ventilação de forma reflexa, para tentar
excreta-lo do organismo (REECE, 2017).
No nível periférico existem
vários receptores que influenciam no controle da respiração entre eles os mecanorreceptores,
os barorreceptores e os quimiorreceptores. Os mecanorreceptores estão presentes
nos músculos, pulmões e vias aéreas e os quimiorreceptores na artéria e na
carótida.
Segundo Reece (2017), os mecanorreceptores localizados nos pulmões quando são
estimulados pelo estiramento durante a inflação dos mesmos transmitem impulsos
via nervo vago para o grupo respiratório dorsal (GRD) sendo muito importantes
para a terminação da inspiração. Esse reflexo segundo o autor é chamado de reflexo
de Hering-Breuer.
Reece (2017) aponta a existência
de receptores de estiramento dos feixes musculares, encontrados nos músculos
estriados, que controlam a força de contração dos músculos respiratórios.
Segundo Klein (2014) o diafragma possui poucos receptores musculares, mas os
músculos intercostais são bem providos de órgãos tendinosos e feixes
musculares. Segundo autor os receptores musculares controlam a força de
contração dos músculos respiratórios e a ajustam quando a ventilação sofre
impedimento, como ocorre com a obstrução das vias aéreas.
Os barorreceptores estão localizados
nos seios aórticos e carotídeos e são sensíveis ao aumento e diminuição da
pressão arterial e capazes de modificar a respiração (REECE, 2107). O autor informa que a elevação da pressão
arterial gera impulsos inibitórios para o centro respiratório e a frequência
respiratória diminui permitindo reduzir o retorno de sangue ao coração. Por
outro lado a pressão arterial reduzida induzirá o aumento das descargas
enviadas para o centro respiratório e consequente aumento da frequência
respiratória e maior fluxo de sangue para o coração. O aumento ou diminuição da
frequência respiratória aumenta ou diminui o número de vezes que ocorre
expansão do tórax e a natural pressão negativa do espaço mediastino que garante
maior ou menor retorno de sangue ao coração.
Os quimiorreceptores existentes no nível periférico recebem
denominação de corpos aórticos e carotídeos segundo Klein (2014) ou glomos
aórticos e carotídeos segundo Reece (2017). Os autores indicam a localização destes
quimiorreceptores próximos à bifurcação das artérias carotídeas interna e
externa, ou ao redor do arco aórtico. Esses receptores periféricos têm a
capacidade de monitorar o O2, CO2 e a concentração do íon
H+ (Ph). Eles fornecem uma descarga tônica para a respiração quando
esta é normal. Conforme a composição do sangue sai do normal, as alterações nas
tensões arteriais de CO2 (PaCO2) e de Oxigênio (PaO2),
e no pH, produzem grandes mudanças na ventilação (KLEIN, 2014).
Os quimiorreceptores periféricos, diferentemente dos
centrais, são capazes de perceber a PaO2 além da PaCO2 e
o pH (KLEIN, 2014; REECE, 2017).
Klein (2014) afirma que os corpos aórticos são supridos
pelo nervo vago parecem ser mais ativos no feto e ter pouca importância no adulto.
Já os corpos carotídeos possuem uma alta
vascularização e são supridos por ramos do nervo glossofaríngeo. Segundo o
autor quando os corpos carotídeos são perfundidos por sangue com baixa PaO2
ou alta PaCO2 ou pH baixo, a frequência de descargas nas fibras
aferentes nervosas do corpo carotídeo aumenta refletindo de modo linear no
aumento da ventilação. Os receptores periféricos são essenciais na tentativa de
recuperação nos quadros de hipóxia quando a acidose induz a hiperventilação, já a alcalose, por sua vez, se
presente, induz uma hipoventilação.
Um outro tipo de receptor periférico é denominado receptor
irritante e acredita-se que esses receptores irritantes, ou receptores de
estiramento de adaptação rápida, sejam terminações nervosas mielinizadas que se
ramificam entre as células epiteliais da laringe, traqueia, grandes brônquios e
vias aéreas intrapulmonares. Eles são estimulados pela deformação mecânica nas
vias aéreas, assim como gases irritantes, acúmulo de muco e a liberação de
histamina e provocam a tosse, secreção de muco, broncoconstrição, respiração
rápida e superficial (hiperpnéia). Todas essas respostas tem efeitos protetivos
para eliminar materiais irritantes do sistema respiratório (KLEIN, 2014).
Outro tipo de receptor
denominado receptor justacapilar, assim chamado pois se localiza dentro do
interstício pulmonar próximo aos capilares pulmonares. Os receptores
justacapilares têm as informações transportadas pelas fibras C e quando
ativados provocam o aumento na frequência respiratória chamada de taquipneia normalmente
associada com doenças vasculares, alérgicas ou infecciosas (Klein,
2014).
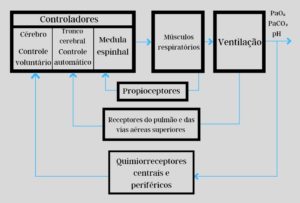
Imagem 2 – Esquema de controle por feedback da regulação da respiração. Legenda: Adaptado de Cunnigham – Tratado de fisiologia veterinária. 5. ed. Rio de Janeiro: Elsevier, 2014.
Além
dos receptores periféricos localizados no pulmões existem receptores
localizados nas vias aéreas superiores. A estimulação dos receptores na
cavidade nasal provoca o ato de fungar e de espirrar, ao passo que aqueles
localizados na laringe e faringe pode causar tosse, apneia e broncoconstrição
(Klein, 2014)
Controle da respiração nas aves
As aves possuem um sistema
respiratório bastante diferente dos mamíferos e conta com sacos aéreos em sua
estrutura e ausência de um músculo diafragma e nervoso frênico. Segundo Ludders (2017) a existência de um centro
de regulação da respiração no cérebro das aves ainda não foi demonstrado de
modo inequívoco, mas à semelhança de mamíferos, o padrão gerador central parece
estar localizado na ponte e no bulbo. O autor informa que além do controle
central, também parece existir o controle químico da respiração através de
quimiorreceptores, além da ação de mecanorreceptores e termorreceptores
extrapulmonares.
Em relação aos
quimiorreceptores, além daqueles extrapulmonares, as aves possuem um grupo que
não está presente nos mamíferos e que são chamados de quimiorreceptores
intrapulmonares (QIP) muito sensíveis ao dióxido de carbono (PCO2) e
insensíveis à hipóxia (Baixa PO2). Estes afetam o volume e
frequência da respiração (LUDDERS, 2017).
Também existem mecanorreceptores
localizados nos sacos aéreos e tecidos adjacentes das aves que são sensíveis a
inflação do sistema respiratório, mas insensíveis a hipóxia e a hipercapnia. E
os barorreceptores arteriais e carotídeos que também influenciam na regulação
da respiração. Quando ocorre aumento da pressão arterial uma hipoventilação é
provocada e quando detecta uma diminuição da pressão arterial, uma
hiperventilação é provocada (LUDDERS, 2017).
Os termorreceptores participam
na regulação da respiração das aves de forma muito ativa para permitir a termorregulação
desses animais. Eles estão localizados na parte central e periférica das aves,
porém as informações são processadas no hipotálamo. A principal via de perda de
calor nas aves vem da evaporação na
respiração, e o sistema respiratório precisa equilibrar a troca gasosa com a
evaporação do calor. Para isso as aves possuem um mecanismo interessante para
fazer o equilíbrio chamado polipnéia, que serão melhor descritos no texto de
termorregulação, mas basicamente aumenta a ventilação do espaço morto, que
aumenta a evaporação, resfriando o corpo da ave, porém sem que haja uma hiperventilação
no nível dos parabrônquios que são a sede de troca gasosas nas aves. (LUDDERS,
2016)
Referências bibliográficas
REECE, W. O. Regulação da respiração. In: REECE, W.O. Dukes – Fisiologia dos animais domésticos. 13ª ed. Rio de Janeiro: Guanabara Koogan, 2016. Não paginado. pt. 4, cap. 24.
KLEIN, B. G. Controle da ventilação . In: KLEIN, B. G. Cunnigham:tratado de fisiologia veterinária. 5. ed. Rio de Janeiro: Elsevier, 2014. Não paginado. cap. 49.
LUDDERS, J. W. Respiração das aves. In: REECE, W. O. Dukes:Fisiologia dos animais domésticos. 13. ed. Rio de Janeiro: Guanabara Koogan, 2016. Não paginado. pt. 4, cap. 26.